Quantitative Immunity in Plants
When attacked by a pathogen, plants are able to set up defense mechanisms that will slow down more or less effectively, and sometimes stop the infection completely. These inducible defenses include, for example, the production of molecules with antimicrobial properties, such as molecules of the glucosinolate and isoflavone family. A plant defense mechanism called hypersensitive response relies on the rapid self-destruction of plant cells bordering the point of infection (Fig. 1). By radically modifying the pathogen's environment, the hypersensitive response can completely inhibit the infection and stop the disease completely. This is called complete resistance or qualitative resistance. The hypersensitive response is particularly effective against biotrophic pathogens, which can only complete their infectious cycle in a living host (Fig. 1). Moreover, the triggering of the hypersensitive response requires the recognition of specific molecules of certain pathogen genotypes by dedicated plant receptor proteins. This form of defence is thus controlled by a more or less elaborate key-lock mechanism that is reconstituted only when plant-pathogen pairs with compatible keys and locks are present (Fig. 2). Varietal selection is often aimed at favouring plants capable of recognising genotypes of pathogens common in agricultural environments. Minor variations in the "molecular key" carried by the pathogen may nevertheless render the plant detection system inoperative (pathogen evading resistance). On the other hand, many pathogens are able to complete their infectious cycle in dead host tissue, and necrotrophic agents will even actively induce host cell death to complete their cycle (Fig. 1). Consequently, the hypersensitive response and the resulting complete resistance remain exceptional phenomena in nature.

Figure 1: Some plant disease symptoms indicative of the molecular mechanisms governing plant-pathogen interaction. (A) Typical hypersensitive reaction (dead cells surrounded by a yellowish halo) on a leaf of A. thaliana leading to the confinement of pathogenic bacteria in a limited area of the leaf, then to their destruction. (B) Young sunflower seedling infected with the downy mildew oomycete Plasmopara halstedii, forming white sporangia on the surface. Infected tissues remain alive and green in response to this biotrophic pathogen. (C) Sunflower plant infected with the white rot fungus Sclerotinia sclerotiorum. The infected plant organs die rapidly, producing brownish lesions characteristic of a necrotrophic interaction
In the absence of qualitative resistance, a phenotype of partial or incomplete resistance is often observed, resulting in a reduction of symptoms rather than absence of disease on a given plant. At the level of plant populations, this phenomenon results in a continuous distribution of resistance levels and is referred to as quantitative disease resistance (QDR, Fig. 2). The emergence of a qualitative or quantitative resistance phenotype depends on the genotype of the pathogen and the plant genotype, according to the molecular key lock model mentioned above, but also on environmental conditions (soil and climate conditions and other microorganisms present). Thus, understanding the nature and regulation of the biological processes that contribute to the quantitative resistance phenotype is essential for optimizing the durability of plant resistance in the face of the evolutionary power of pathogen populations and variations in environmental conditions.
The genetic architecture of the resistance trait studied is often rich in information on the evolutionary and functional constraints that weigh on plant immunity, but is sometimes difficult to approach. The mapping of quantitative resistance loci typically leads to the identification of multiple quantitative trait loci (QTLs) of low to moderate effect, explaining between 5 and 30% of the phenotypic variance. In theory, qualitative resistance is therefore distinguished by the possibility of obtaining complete resistance by the introduction of a single gene (known as "resistance" or "R" gene) into a sensitive genetic background, whereas the introduction of a quantitative resistance gene will only confer partial resistance. In practice, however, some R genes may only confer partial resistance depending on the allele considered, the environmental conditions or the genotype of the pathogen under study (Fig. 2). Conversely, a quantitative resistance phenotype may appear binary (resistant/sensitive typical of qualitative resistance) if the plant population under study lacks diversity, if environmental conditions erase phenotypic differences, or if the precision and resolution of the phenotyping methodology is insufficient. Functional characterization of resistance QTLs therefore requires robust and accurate technologies to (i) map quantitative resistance loci on the plant genome and (ii) assess the phenotypic contribution of a gene or allele to the resistance locus.
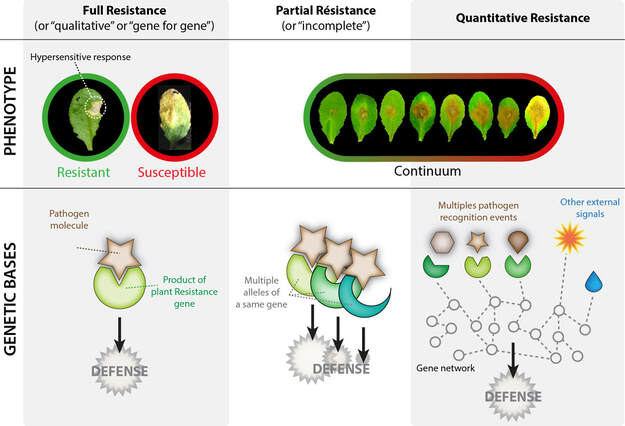
Figure 2. Different forms of plant resistance. These are characterized by the distribution of the resistance phenotype within a host population (resistance-sensitive binary or continuum), as well as by the architecture of the genetic bases involved. Molecules produced by the pathogen are shown in brown, molecules produced by the plant are shown in green. In the gene network, each circle represents a gene of the plant. The dotted lines represent interactions between the elements of the network. The "other external signals" correspond to signals from the environment that are independent of the pathogen, such as light intensity, humidity, etc. The "other external signals" correspond to signals from the environment that are independent of the pathogen, such as light intensity, humidity, etc.
Recommended reading:
- Genome evolution in filamentous plant pathogens: why bigger can be better. www.nature.com/articles/nrmicro2790
- Resistance to phytopathogens e tutti quanti: placing plant quantitative disease resistance on the map. bsppjournals.onlinelibrary.wiley.com/doi/full/10.1111/mpp.12138
- Advances on plant-pathogen interactions from molecular toward systems biology perspectives. onlinelibrary.wiley.com/doi/full/10.1111/tpj.13429
- Silent control: microbial plant pathogens evade host immunity without coding sequence changes. academic.oup.com/femsre/advance-article/doi/10.1093/femsre/fuab002/6095737
Sclerotinia, the hard-hitting mold
The white and stem mold pathogen Sclerotinia sclerotiorum is a generalist fungal pathogen, infecting a broad range of host species (>400) in nature. Its name comes from the from Ancient Greek σκληρός (sklērós) which means “hard", by reference to the hard and melanized resting structures called sclerotia that it forms. This greek word also means "tough”, which accurately reflects how Sclerotinia behave with plants. It is among the most devastating plant pathogens worldwide and causes disease on many crops including soybean, rapeseed, sunflower and most vegetables. Solutions for the genetic control of the disease are very limited in most crops. S. Sclerotiorum naturally infects wild and cultivated Brassica species, including the model plant Arabidopsis thaliana which shows quantitative disease resistance when challenged by Sclerotinia.
A majority of studies on plant interactions with fungal pathogens over the last years have focused on tightly coupled host-pathogen interactions. For instance the powdery mildew fungus Blumeria graminis, the cereal rust fungi of the Puccinia spp., and the corn smut fungus Ustilago maydis are among the most studied fungal pathogens and are obligate biotrophic pathogens restricted to a single host genus. Such pairwise interactions only represents a fraction of plant-fungal pathogen interactions encountered in nature and a number of broad host range fungal pathogens also are major threats for food security. Understanding how broad host range pathogens successfully infect multiple plant lineages is a major challenge in plant pathology.
Among Leotiomycete, the grey mold fungus Botrytis cinerea and the white mold fungus Sclerotinia sclerotiorum stand out for having a remarkably broad host range, encompassing over 200 species. Each of these pathogens causes yearly several hundred millions of US dollars crop losses worldwide. They are considered as typical necrotrophs, secreting an arsenal of cell wall-degrading enzymes and toxins to kill host cells and derive energy. Host plants typically exhibit quantitative disease resistance (QDR) to B. cinerea and S. sclerotiorum, leading to a reduction rather than absence of disease. How these generalist fungal pathogens cause disease and what are the genetic bases of plant QDR is still poorly understood and a major focus of our research.
A majority of studies on plant interactions with fungal pathogens over the last years have focused on tightly coupled host-pathogen interactions. For instance the powdery mildew fungus Blumeria graminis, the cereal rust fungi of the Puccinia spp., and the corn smut fungus Ustilago maydis are among the most studied fungal pathogens and are obligate biotrophic pathogens restricted to a single host genus. Such pairwise interactions only represents a fraction of plant-fungal pathogen interactions encountered in nature and a number of broad host range fungal pathogens also are major threats for food security. Understanding how broad host range pathogens successfully infect multiple plant lineages is a major challenge in plant pathology.
Among Leotiomycete, the grey mold fungus Botrytis cinerea and the white mold fungus Sclerotinia sclerotiorum stand out for having a remarkably broad host range, encompassing over 200 species. Each of these pathogens causes yearly several hundred millions of US dollars crop losses worldwide. They are considered as typical necrotrophs, secreting an arsenal of cell wall-degrading enzymes and toxins to kill host cells and derive energy. Host plants typically exhibit quantitative disease resistance (QDR) to B. cinerea and S. sclerotiorum, leading to a reduction rather than absence of disease. How these generalist fungal pathogens cause disease and what are the genetic bases of plant QDR is still poorly understood and a major focus of our research.
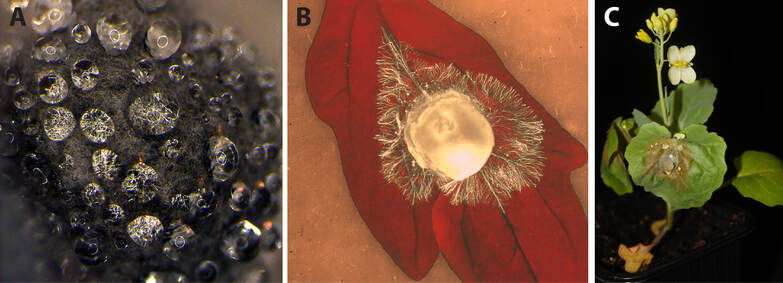
Figure 3. The plant pathogenic fungus Sclerotinia sclerotiorum.
Recommended reading:
- Emerging Trends in Molecular Interactions between Plants and the Broad Host Range Fungal Pathogens Botrytis cinerea and Sclerotinia sclerotiorum. www.frontiersin.org/articles/10.3389/fpls.2016.00422/full
- Shifts in diversification rates and host jump frequencies shaped the diversity of host range among Sclerotiniaceae fungal plant pathogens. onlinelibrary.wiley.com/doi/full/10.1111/mec.14523
- Codon optimization underpins generalist parasitism in fungi. elifesciences.org/articles/22472
- Intercellular cooperation in a fungal plant pathogen facilitates host colonization. www.pnas.org/content/116/8/3193.long
